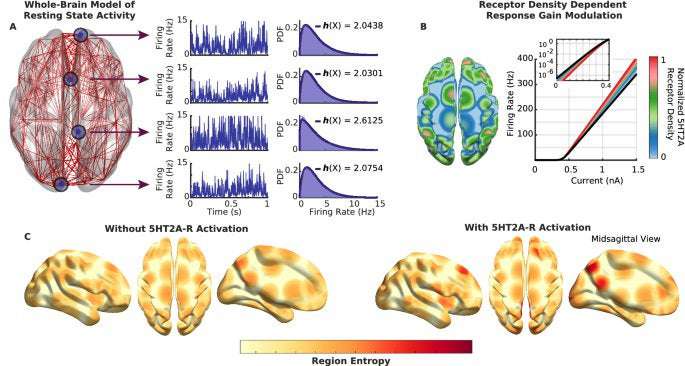
In this study we investigated the brain entropy changes induced by serotonergic psychedelics by simulating whole-brain resting state activity with and without 5HT2A-R activation. In contrast to empirical studies, which usually only have access to coarse-grained fMRI or M/EEG data, our approach allows us to study firing rates of brain regions and hence to directly assess the effect of 5HT2A-R agonism at the level of neural population activity.
In agreement with the Entropic Brain Hypothesis14, the model shows a significant increase of global brain entropy, albeit in a highly inhomogeneous and non-linear reconfiguration. The diversity of effects stresses the importance of extending the scope of the EBH from a simple level-based approach towards a multi-dimensional perspective, which might better characterise the richness of 5HT2A-R activation effect both on the brain and consciousness.
We propose a possible mechanism in the sense that the average increase in neural entropy induced by serotonergic psychedelic drugs can be explained (in statistical sense) in terms of quantities that provide a neurobiological interpretation about how this phenomena could occur in the brain. These results have important consequences for our understanding of consciousness, neurodynamics, and the psychedelic state. In the following, we highlight some of those implications, and propose possibilities for future work.
5HT2A-R-induced entropy changes are regionally heterogeneous
Our main result in this study is a mechanistic explanation of the main prediction of the EBH that serotonergic psychedelics increase the entropy of brain activity. However, one of the main takeaways is that this overall increase is tied to a spatially heterogeneous reconfiguration, rather than a globally consistent increase, in entropy.
Our simulation results show that 5HT2A-R activation triggers an entropy increase in sensory areas, as observed in fusiform and olfactory cortices, as well as the primary visual RSN and occipital regions (Supp. Fig. 1). This agrees with the increased perceptual ‘bandwidth’ that characterises the psychedelic state29, as higher entropy might be related to richer perceptual experience in a given moment of consciousness—potentially related to reduced gating. As a matter of fact, all the primary visual RSN regions (with the exception of the lingual area) are part of the high-strength group, suggesting that the effect of psychedelics on perceptual experience might be directly related to 5HT2A-R density in those regions. The localisation of the entropy increases may relate to domain-specific changes in consciousness, which could be interpreted as consistent with a recent dimensions of consciousness characterisation of the psychedelic experience30.
Comparison to in vivo experiments with psychedelic drugs
Throughout the paper, we have focused on one particular signature of 5HT2A-R agonists on the brain—a global increase in average entropy. But much more is known about the effect of psychedelics on the brain, and studying these more nuanced effects is key to understanding the rich phenomenology of the psychedelic state. In this section we deepen this connection by providing a more detailed comparison between the behaviour of the model and experimental results with psychedelic drugs.
Our findings are in agreement with earlier studies where the effect of psychedelic drugs on the topographical distribution of entropy-related measures was correlated with subjective effects. For example, Schartner et al.9 studied the effect of LSD, psilocybin, and ketamine on the entropy rate of binarized MEG signals, and found localised increases in entropy rate on occipital and parietal regions. Similarly, Lebedev et al.10 analysed the sample entropy of fMRI recordings of subjects under the effect of LSD, finding localized increases on frontoparietal, medial occipital, posterior and dorsal cingulate regions. Many of those regions showed a consistent increase of their entropy with 5HT2A-R agonism in our study (Fig. 2A).
Moreover, many of those regions actually belong to the high connectivity strength group (c.f. Fig. 3), which suggests that their entropy increase in experimental data might be directly related to the high 5HT2A-R density in those regions. We speculate that regions of the high-strength group may systematically exhibit significant differences in local entropy on future experiments. Together, these findings support the conclusion that the DMF model, once optimised, can reproduce not only functional connectivity18, but also some of the most salient localised entropy increases found on in vivo human studies up to date.
Also, on a more fundamental level, our findings suggest that psychedelics disrupt the functional organisation of the brain with an especially focal and pronounced action on highly anatomically connected brain regions. In fact, experimental and modelling evidence points towards a key role of connector hubs on psychedelic and psychopatological states31,32. Perturbing the activity in these specific regions (via 5HT2A-R agonism) could have particularly profound implications for the regularity of brain activity and the quality of conscious experience. However, sub cortical regions may also play an important role on the modulation of brain activity, conscious experience, and the psychedelic state33.
On a separate line of inquiry, there is strong evidence associating the DMN to high-level cognitive functions, including the sense of space, time and recognition of (self) boundaries. Disruptions to the DMN have been linked to fundamental changes in experience, such as ego dissolution14,34. Our simulation results show that 5HT2A-R activation increases the entropy of all DMN regions (with the exception of the angular cortex), which is consistent with the reported decrease in the DMN network integrity31 induced by psychedelic drugs. In contrast, low-level motor functions such as motor regulation remain largely unaffected during the psychedelic state35, which is consistent with the modest entropy changes observed in the lingual and superior motor areas.
Finally, it is worth noting that both angular regions showed particularly important decreases in local entropy, specially in the left hemisphere (Fig. 2A). Damage to this region is associated with impairments on language processing, and electrical stimulation can induce out-of-body experiences36 (both experiences can feature within psychedelic states37). Since both structural damage and electrical stimulation can be related to entropy reduction (e.g. by stimulus-driven synchronisation), these experimental findings are consistent with the strong entropy decrease in angular regions predicted by the model.
We speculate that the reduced entropy observed on the angular region induced by 5HT2A-R agonism could be related to a smaller local bandwidth, which in turn might impair the multi-modal and integrative functions of this region36.
The approach employed in this work presents certain limitations related to several aspects of the simulation and analysis. Acknowledging and understanding these limitations can help us extend and improve our approach, while introducing new questions in the field of psychedelic computational neuroscience.
To our knowledge, the DMF model is the only whole-brain model that implements neuromodulation and is capable of reproducing neuroimaging data in the placebo and psychedelic states. Nonetheless, it makes some important simplifications that are worth discussing. At the network level, the DTI-based connectome used here is known to be incomplete, thus improvements could be made to the model parameters of brain connectivity38. At the dynamical level, the DMF model models neuronal populations as perfectly homogeneous within a given region, and it is known that finer-grained local structure of certain brain regions is likely to be key to explaining certain subjective effects of the psychedelic state (e.g. lattice structure in the visual cortex and geometric visual hallucinations39). Additionally, the version of the DMF model used here only considers 5HT2A-R agonism, while classic serotonergic psychedelic drugs also have high binding affinities for other receptors (e.g. in the case of LSD, the D1 and D2 dopamine receptors40).
These simplifications do not prevent the model from reproducing statistical features of brain signals under the placebo and 5HT2A-R conditions, but could result in an inability to reproduce finer aspects of the dynamics of the whole-brain activity in these conditions. Extending the model to reproduce other dynamical signatures of psychedelics (like alpha suppression 8 or reduced directed functional connectivity 41) constitutes a natural extension of this work, with the recent example of Kringelbach et al. 42, where an extension of the DMF can reproduce dynamical features of the brain in placebo and in the psychedelic state.
Another potentially fruitful line of future work involves making more detailed comparisons with in vivo psychedelic neuroimaging data, and, potentially, subjective experience reports. For example, one natural option would be to use forward models of fMRI43 or M/EEG44 to bridge between the firing rates produced by the DMF model and other data modalities, to produce simulated data that is more directly comparable with available empirical data.
Another exciting possibility is to explore model parameters to examine potential non-linearities and their implications for different relevant aspects of brain function. For example, there are reasons to believe that the dose-response relationship is non-linear for psychedelics and that over a certain threshold dosage (level of 5HT2A-R stimulation) new subjective and global brain function properties can appear45.
Most interestingly, a potentially very useful extension of this work would be to include individual subject-level connectome and receptor data to build personalised models of response to psychedelic drugs. This would enable a much more comprehensive modelling framework, capable of correlating structural brain features with subjective experience reports. Such a framework could potentially make individualised predictions of the action of serotonergic psychedelics on specific individuals, aiding patient stratification and treatment customisation2.
Finally, it is worth noting that all our analyses here are based on the univariate statistics of individual brain regions, not including any correlation or information flow between them. However, it is known that some high-level subjective effects of psychedelics (such as complex imagery7 and ego dissolution34) are related to network, as opposed to single region, dynamics. Therefore, building a richer statistical description of the brain’s dynamics using recent information-theoretic tools (such as multivariate extensions of mutual information28 or Integrated Information Theory, IIT46) remains an exciting open problem. In fact, recent attempts of unifying IIT and the EBH in a single framework to understand the effects of psychedelic drugs47 are helping to bridge the gap between the univariate EBH and the multivariate IIT.
In this paper we have provided the first mechanistic explanation of the neural entropy increase elicited by psychedelic drugs, using a whole-brain dynamical model with 5HT2A-R neuromodulation. Furthermore, we built a simple linear model able to predict a region’s relative change in entropy from its local 5HT2A-R density and topological properties, showing that, somewhat paradoxically, at a whole-brain level receptor density is a poor predictor of 5HT2A-R activation effect.
Key to developing this predictive model was a three-way partition of brain regions according to their connectivity strength, suggesting a differentiated action mechanism of 5HT2A-R agonists that depends on the local topology of brain regions. In summary, our results suggest that the local changes in entropy, as well as the global entropy increase, induced by 5HT2A-R activation can be explained from a region-specific interplay between structural connectivity and receptor density. Finally, controlled experiments with null network models confirm that receptor density and connectivity strength are not only necessary, but also sufficient, to explain the entropic effects of 5HT2A-R activation.
The spatially heterogeneous, complex nature of the observed effects of 5HT2A-R activation opens a challenging problem for understanding the clinical and scientific relevance of psychedelic drugs and their entropic effect. Furthermore, it stresses the necessity of moving beyond the current unidimensional approach to consciousness to a multi-dimensional one, that better captures the phenomenological and neurodynamical richness of psychedelic state.
PolyMorpheusPervert on March 15th, 2021 at 07:29 UTC »
So basically the "Doors of Perception" by Aldous Huxley. Only around 70 years later did science prove him correct(ish).
PaladinsLover69 on March 15th, 2021 at 05:59 UTC »
Holy cow...this is cool. I want to be in a study. How do I do that?
mvea on March 15th, 2021 at 02:42 UTC »
I’ve linked to the open access full-text source journal article in the post. The citation of the article is here:
Herzog, R., Mediano, P.A.M., Rosas, F.E. et al.
A mechanistic model of the neural entropy increase elicited by psychedelic drugs.
Sci Rep 10, 17725 (2020).
Published 20 October 2020
DOI: https://doi.org/10.1038/s41598-020-74060-6
Abstract
Psychedelic drugs, including lysergic acid diethylamide and other agonists of the serotonin 2A receptor (5HT2A-R), induce drastic changes in subjective experience, and provide a unique opportunity to study the neurobiological basis of consciousness. One of the most notable neurophysiological signatures of psychedelics, increased entropy in spontaneous neural activity, is thought to be of relevance to the psychedelic experience, mediating both acute alterations in consciousness and long-term effects. However, no clear mechanistic explanation for this entropy increase has been put forward so far. We sought to do this here by building upon a recent whole-brain model of serotonergic neuromodulation, to study the entropic effects of 5HT2A-R activation. Our results reproduce the overall entropy increase observed in previous experiments in vivo, providing the first model-based explanation for this phenomenon. We also found that entropy changes were not uniform across the brain: entropy increased in some regions and decreased in others, suggesting a topographical reconfiguration mediated by 5HT2A-R activation. Interestingly, at the whole-brain level, this reconfiguration was not well explained by 5HT2A-R density, but related closely to the topological properties of the brain’s anatomical connectivity. These results help us understand the mechanisms underlying the psychedelic state and, more generally, the pharmacological modulation of whole-brain activity.
The reference to perceptual bandwidth is from here:
This agrees with the increased perceptual ‘bandwidth’ that characterises the psychedelic state29, as higher entropy might be related to richer perceptual experience in a given moment of consciousness—potentially related to reduced gating.